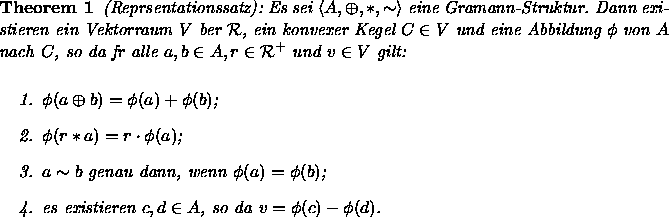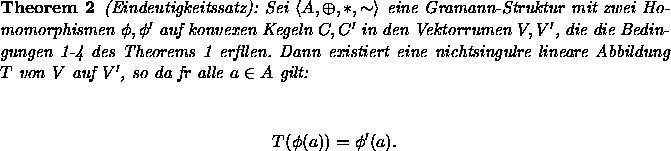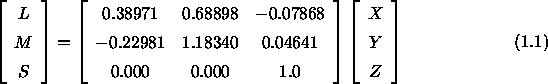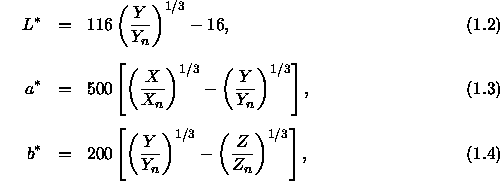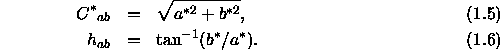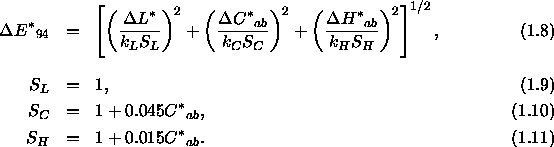Ausgangspunkt für die Farbwahrnehmung sind die ihr zugrundeliegenden physikalischen Farbreize, also diejenigen elektromagnetischen Strahlungen im Bereich zwischen 380 und 780 nm, die in das Auge gelangen und dort spezielle Sinneszellen erregen, so daß eine Farbempfindung entsteht. Ein Farbreiz kann auch als Punkt in einem unendlich-dimensionalen Raum aufgefaßt werden, in dem jede Achse die Intensität einer bestimmten Wellenlänge (beispielsweise in Photonen pro Sekunde) darstellt (siehe Krantz, 1975a). Da sich unendlich-dimensionale Räume nur schwer darstellen lassen, werden Farbreize meist durch ihre spektrale Strahlungsverteilung charakterisiert, die angibt, wie sich die Strahlungsleistung auf die einzelnen Regionen des Spektrums verteilt.
Spezielle Rezeptoren auf der Retina des Auges absorbieren einen Teil der Farbreize, wodurch eine chemische Reaktion ausgelöst wird, die im Zerfall von verschiedenen Arten von Sehpurpur besteht und die anschließend in eine Nervenerregung umgesetzt wird. Dabei bilden ein achromatischer und drei chromatische Mechanismen![]() unterschiedlich gewichtete Summen der Intensitäten bei den verschiedenen Wellenlängen, wobei nicht die gesamte Information über die Intensitäten für alle Wellenlängen erhalten bleibt.
unterschiedlich gewichtete Summen der Intensitäten bei den verschiedenen Wellenlängen, wobei nicht die gesamte Information über die Intensitäten für alle Wellenlängen erhalten bleibt.
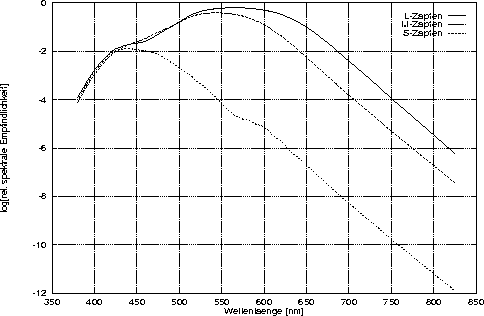
Bei den Rezeptorarten unterscheidet man zwischen den extrafoveal gelegenen Stäbchen und den vor allem auf der fovea centralis angesiedelten Zapfen, von denen sich drei verschiedene Typen anhand ihrer Farbpigmente identifizieren lassen. In Abbildung 1.1 sind die von Smith und Pokorny (1975) durch Farbabgleiche von Dichromaten bestimmten spektralen Empfindlichkeiten dieser drei Zapfentypen dargestellt, deren Maximum im langwelligen (L-System), mittleren (M-System) oder kurzwelligen (S-System) Bereich des sichtbaren Spektrums liegt. Baylor, Nunn und Schnapf (1987) finden bei direkten elektrophysiologischen Messungen (sogenannten suction electrode recordings) an der Retina von Makaken sehr ähnliche Funktionen, die auch mit den von Schnapf, Kraft und Baylor (1987) an Zapfen aus der menschlichen Retina bestimmten Rezeptorempfindlichkeits-Funktionen übereinstimmen. Bei Betrachtung von Abbildung 1.1 zeigt sich, daß sich die Absorptionsspektren aller drei auch als Farbkanäle bezeichneten sensorischen Systeme bei jeder Wellenlänge überlappen. Dies bedeutet, daß beliebige Spektren immer alle drei Systeme - wenn auch in unterschiedlichem Ausmaß - aktivieren.
Relative spektrale Empfindlichkeiten der Zapfentypen: Hier sind die logarithmierten relativen spektralen Empfindlichkeiten nach Smith und Pokorny (1975) für die L-, M- und S-Zapfen dargestellt. Sie dienen als Schätzung der Absorptionsfunktion der jeweiligen Zapfentypen ohne Berücksichtigung des Einflusses des Augenmediums.
Die spektralen Hellempfindlichkeitskurven für das Tagessehen (siehe z.B. Vos, 1978) und das Nachtsehen (nach CIE, 1951) beschreiben die Empfindlichkeit des visuellen Systems und insbesondere des Stäbchen-Mechanismus für die Wahrnehmung monochromatischer Reize in Abhängigkeit von deren Wellenlänge. Die von Kraft, Schneeweis und Schnapf (1993) durch direkte elektrophysiologische Messungen an Stäbchen in der Retina des Menschen ermittelten Funktionen stimmen ebenfalls sehr gut mit den oben genannten, durch Farbabgleiche ermittelten, überein.
Ausführlichere Darstellungen der physiologischen Grundlagen des Farbensehens sind beispielsweise bei Brindley (1970), Pugh (1988), Wandell (1995) oder Kaiser und Boynton (1996) zu finden. In der vorliegenden Arbeit liegt das Interesse jedoch mehr auf der Wahrnehmung der elektromagnetischen Schwingungen im sichtbaren Bereich, die man als Farbe bezeichnet. Die DIN-Norm 5033 legt dazu folgende Definition fest:
Farbe ist diejenige Gesichtsempfindung eines dem Auge strukturlos erscheinenden Teiles des Gesichtsfeldes, durch die sich dieser Teil bei einäugiger Beobachtung mit unbewegtem Auge von einem gleichzeitig gesehenen, ebenfalls strukturlosen angrenzenden Bezirk allein unterscheiden kann. (DIN 5033, Blatt 1)
Die von Objekten unserer Umgebung ausgelöste Farbempfindung, die auch als Objektfarbe bezeichnet wird, hängt sowohl vom Spektrum der Beleuchtung als auch von der vom beleuchteten Objekt reflektierten Strahlung ab. Außerdem übt die Zusammensetzung des zeitlichen und räumlichen Kontextes einen deutlichen Einfluß auf den von einem Reiz hervorgerufenen Eindruck aus, der im Abschnitt 1.3 eingehend dargestellt wird. Zeitliche Kontexteffekte, die auch als Adaptationseffekte bezeichnet werden, zeigen sich in der Veränderung des von einem Reiz hervorgerufenen Farbeindrucks durch vorheriges länger andauerndes Fixieren eines andersfarbigen Reizes.
Ein und dasselbe Objekt erzeugt bei unterschiedlicher Beleuchtung zwar verschiedene Farbreize, das visuelle System sorgt aber dafür, daß dem Beobachter die Farbe des Objekts dennoch konstant erscheint. Eine bekannte Theorie zur Erklärung dieses als Farbkonstanz bezeichneten Phänomens ist beispielsweise diejenige der Normierung mit Durchschnitt (D'Zmura & Lennie, 1986; Dannemiller, 1989): Nach dieser Ansicht ist der Durchschnitt aller Reflektanzen in jeder natürlichen Szene für jede Wellenlänge gleich und wird als farblos wahrgenommen, so daß sich die Farbe eines Objektes unabhängig von der Beleuchtung schätzen läßt. Diese Farbe kann auf unterschiedliche Weise beschrieben werden, wie im nächsten Abschnitt zu sehen ist.
Man kann Farben bezüglich der drei direkt wahrnehmbaren Attribute Helligkeit (brightness), Farbton (hue) und Sättigung (saturation) beschreiben, die beispielsweise bei Wyszecki und Stiles (1982, S. 487) oder im International Lighting Vocabulary (CIE, 1987) definiert sind. Verschiedene Modelle zur Kennzeichnung des farblichen Aussehens (color appearance models, siehe Fairchild, 1998) sagen einige oder alle dieser Farbattribute vorher. Man kann Farben aber auch auf nicht direkt wahrnehmbaren Dimensionen charakterisieren.
Die von einem Farbreiz hervorgerufene Farbempfindung ist nicht direkt proportional zu dessen Intensität und läßt sich durch die Angabe von nur drei Kennwerten repräsentieren: Zu einer beliebigen Farbe existiert immer eine gleich aussehende eigentliche![]() oder uneigentliche
oder uneigentliche![]() Mischung von drei Primärreize genannten spektralen Energieverteilungen, durch deren Intensitäten, die auch als Farbwerte bezeichnet werden, diese Farbe beschrieben werden kann. Die Versuchspersonen von Stiles und Burch (1959) stellen durch solche Mischungen eine Farbgleichheit zu vorgegebenen monochromatischen Reizen verschiedener Wellenlänge
Mischung von drei Primärreize genannten spektralen Energieverteilungen, durch deren Intensitäten, die auch als Farbwerte bezeichnet werden, diese Farbe beschrieben werden kann. Die Versuchspersonen von Stiles und Burch (1959) stellen durch solche Mischungen eine Farbgleichheit zu vorgegebenen monochromatischen Reizen verschiedener Wellenlänge ![]() her, woraus sich durch Variieren der Wellenlängen der monochromatischen Reize über den sichtbaren Bereich die spektralen Farbwertkurven
her, woraus sich durch Variieren der Wellenlängen der monochromatischen Reize über den sichtbaren Bereich die spektralen Farbwertkurven ![]() bestimmen lassen. Diese Koeffizienten geben die Intensitäten der drei Primärreize an, die für einen Farbabgleich zu einem monochromatischen Reiz einer bestimmten Wellenlänge erforderlich sind.
bestimmen lassen. Diese Koeffizienten geben die Intensitäten der drei Primärreize an, die für einen Farbabgleich zu einem monochromatischen Reiz einer bestimmten Wellenlänge erforderlich sind.
Bereits Maxwell (1856/1970) weist darauf hin, daß bei der additiven Farbmischung die Wahl der Primärreize weitgehend beliebig ist, so daß ``any other three colors might have been chosen, provided that white resulted from their combination in proper proportions'' (Maxwell, 1856/1970, S. 64). Einzige Voraussetzung ist lineare Unabhängigkeit der Primärreize, was praktisch bedeutet, daß sich keiner der Primärreize aus den beiden anderen ermischen lassen darf. Monochromatische Primärreize werden allerdings bevorzugt, da sich bei diesen die neuen Farbwerte besonders leicht berechnen lassen, wenn andere Primärreize verwendet werden.
Die im vorigen Abschnitt beschriebene numerische Repräsentation von Farben durch ihre Farbwerte soll nun meßtheoretisch betrachtet werden. Auf Graßmann (1853) geht die Darstellung von Farben als Elemente eines dreidimensionalen Vektorraums zurück und Krantz (1975a) axiomatisiert diesen Ansatz im Rahmen der Meßtheorie von Krantz, Luce, Suppes und Tversky (1971). Unendlich-dimensionale Vektoren, die die Farbreize repräsentieren, lassen sich durch Einführung einer geeigneten Äquivalenzrelation in einem Vektorraum geringerer Dimensionalität (bei den Farben beträgt sie drei) repräsentieren. Nun soll dargestellt werden, wie dies möglich ist.
Grundlage der Repräsentation von Farben durch Vektoren bilden zwei Abbildungen und eine Relation: Die Abbildung ![]() bezeichne die Mischung zweier Reize und die Abbildung
bezeichne die Mischung zweier Reize und die Abbildung ![]() bedeute die Steuerung oder Variation der Reizintensität, z.B. durch Verwendung eines Graukeils. Empfundene Farbgleichheit zweier Reize, auch Metamerie genannt, werde schließlich durch die Metamerierelation
bedeute die Steuerung oder Variation der Reizintensität, z.B. durch Verwendung eines Graukeils. Empfundene Farbgleichheit zweier Reize, auch Metamerie genannt, werde schließlich durch die Metamerierelation ![]() ausgedrückt. Bei den beiden Abbildungen handelt es sich um physikalische Veränderungen des Reizes, bei der an dritter Stelle genannten Relation dagegen um die (psychologische) Beurteilung zweier Reize. Krantz (1975a) zeigt nun, daß sich additive Farbmischung als positive Halbgruppe mit Aufhebungseigenschaft und Intensitätsveränderungen als skalare Multiplikation auffassen lassen, wenn die Voraussetzungen einer sogenannten Graßmann-Struktur gegeben sind:
ausgedrückt. Bei den beiden Abbildungen handelt es sich um physikalische Veränderungen des Reizes, bei der an dritter Stelle genannten Relation dagegen um die (psychologische) Beurteilung zweier Reize. Krantz (1975a) zeigt nun, daß sich additive Farbmischung als positive Halbgruppe mit Aufhebungseigenschaft und Intensitätsveränderungen als skalare Multiplikation auffassen lassen, wenn die Voraussetzungen einer sogenannten Graßmann-Struktur gegeben sind:
Axiom 4 beschäftigt sich mit der Verträglichkeit von ![]() und
und ![]() : Die Metamerierelation zwischen zwei Farben soll erhalten bleiben, wenn zu beiden eine jeweils gleiche dritte Farbe hinzugemischt wird. Die Verträglichkeit von
: Die Metamerierelation zwischen zwei Farben soll erhalten bleiben, wenn zu beiden eine jeweils gleiche dritte Farbe hinzugemischt wird. Die Verträglichkeit von ![]() und
und ![]() wird in Axiom 5 gefordert, d.h. die Metamerierelation soll auch gelten, wenn die Intensität zweier anfangs metamerer Farben um den gleichen Betrag erhöht oder verringert wird. Zur Beschreibung der Dimensionalität des Farbensehens ist schließlich folgende Definition wichtig:
wird in Axiom 5 gefordert, d.h. die Metamerierelation soll auch gelten, wenn die Intensität zweier anfangs metamerer Farben um den gleichen Betrag erhöht oder verringert wird. Zur Beschreibung der Dimensionalität des Farbensehens ist schließlich folgende Definition wichtig:
Bei Vorliegen einer m-chromatischen Graßmann-Struktur lassen sich nach dem ersten Teil dieses Axioms bei m+1 Primärreizen immer verschiedene Möglichkeiten finden, sie zu einer bestimmten Farbe zu mischen. Der zweite Teil behauptet, daß es bei m Primärreizen dagegen genau eine Möglichkeit gibt, diese so zu mischen, daß deren Mischung farbgleich zu einer vorgegebenen Farbe ist. Nach dem sogenannten ersten Graßmannsche Gesetz gilt für farbnormale Beobachter die Trichromatizität des Farbensehens, die Farbwahrnehmung ist also dreidimensional.
Die hier zusammengestellten Axiome lassen sich in physikalische und psychologische einteilen: Die Axiome 1 und 2 befassen sich mit den physikalischen Eigenschaften von Farbmischungen, die Axiome 3 bis 6 betreffen die Farbwahrnehmung. Sind die Axiome der Graßmann-Struktur empirisch erfüllt, dann können Farben durch dreidimensionale Vektoren repräsentiert werden, wie die beiden folgenden Theoreme von Krantz (1975a) feststellen:
Die Abbildung ![]() ist ein Homomorphismus der Graßmann-Struktur
ist ein Homomorphismus der Graßmann-Struktur ![]() auf
auf ![]() , wobei C ein konvexer Kegel in V ist. Wegen Eigenschaft 4 handelt es sich bei V um einen minimalen Vektorraum, da sich jedes Element von V als Differenz von Elementen
, wobei C ein konvexer Kegel in V ist. Wegen Eigenschaft 4 handelt es sich bei V um einen minimalen Vektorraum, da sich jedes Element von V als Differenz von Elementen ![]() und
und ![]() aus C erzeugen läßt (siehe Krantz, 1975a).
aus C erzeugen läßt (siehe Krantz, 1975a).
Die Abbildung ![]() ist damit eindeutig bis auf nichtsinguläre lineare Transformationen.
ist damit eindeutig bis auf nichtsinguläre lineare Transformationen.
Wie durch Theorem 1 gezeigt wird, läßt sich die Menge der Farbreize strukturerhaltend in einen Vektorraum abbilden: Nach Drösler (1989) besteht der psychologische Teil des Gedankengangs darin, die Farbreize abzubilden auf einen Vektorraum von Äquivalenzklassen von Reizen; dieser Farbraum kann dann wiederum auf den numerischen Vektorraum ![]() abgebildet werden. Die zulässigen Transformationen T aus Theorem 2 drücken die Beliebigkeit der Auswahl des Tripels an Primärreizen aus, die die Basis des Vektorraums bilden.
abgebildet werden. Die zulässigen Transformationen T aus Theorem 2 drücken die Beliebigkeit der Auswahl des Tripels an Primärreizen aus, die die Basis des Vektorraums bilden.
Nach Krantz (1975a) können alle eindimensionalen Merkmale, die dazu dienen, Farben zu unterscheiden, als Farbcodes bezeichnet werden, beispielsweise auch die in Abschnitt 1.1.1 genannten Farbattribute. Wenn sich zwei Farben in einem ihrer Farbcodes unterscheiden, kann keine Metamerierelation zwischen ihnen bestehen. Umgekehrt können aber Farben mit einem gleichen Farbcode unterschieden werden, wenn Unterschiede in einem anderen Farbcode bestehen. Suppes, Krantz, Luce und Tversky (1989) weisen darauf hin, daß diese Farbcodes sowohl auf der neuronalen bzw. Rezeptorebene (neural codes) bestehen können als auch auf der Wahrnehmungsebene (perceptual codes).
Mausfeld (1998) nennt eine Reihe von Einschränkungen bezüglich der Anwendbarkeit der Graßmann-Struktur auf Farben: Ein wesentlicher Schwachpunkt besteht darin, daß sich damit nur isoliert dargebotene Farben beschreiben lassen, für komplexere Reizkonfigurationen ist sie weniger geeignet. Da die Metamerierelation die einzige psychologische Relation ist, die dem Vektorraum zugrunde liegt, lassen sich die euklidischen Distanzen zwischen Vektoren nicht als Maße der Farbähnlichkeit oder der Unterscheidbarkeit interpretieren. Graßmann-Strukturen erfassen damit nicht die Ausprägung von Farbattributen wie Farbton, Sättigung oder Helligkeit und auch nicht das farbliche Aussehen (color appearance), das vom Kontext und der Interpretation des Beobachters bestimmt wird.
Nun sollen konkrete Realisierungen der im letzten Abschnitt abstrakt eingeführten numerischen Farbrepräsentation vorgestellt werden, die sich durch die Festlegung eines Primäreiz-Tripels ergeben. Zwischen den verschiedenen Farb-Systemen lassen sich die Koordinaten der Farben nach dem in Theorem 2 gegebenen Eindeutigkeitssatz umrechnen. Das ![]() -System (tristimulus color-space) ist ein Koordinatensystem, das auf den drei monochromatischen Primärreizen Rot
-System (tristimulus color-space) ist ein Koordinatensystem, das auf den drei monochromatischen Primärreizen Rot ![]() (700,0 nm), Grün
(700,0 nm), Grün ![]() (546.1 nm) und Blau
(546.1 nm) und Blau ![]() (435.8 nm) basiert. Grundlage bilden die von der CIE (1931) bestimmten Spektralwertkurven, die angeben, welche Mischungen dieser Primärreize farbgleich sind zu einem monochromatischem Reiz bestimmter Wellenlänge.
(435.8 nm) basiert. Grundlage bilden die von der CIE (1931) bestimmten Spektralwertkurven, die angeben, welche Mischungen dieser Primärreize farbgleich sind zu einem monochromatischem Reiz bestimmter Wellenlänge.
Jeder der drei Farben entspricht dabei eine Achse (bzw. ein Basisvektor) des Farbraumes und die Einheitsebene (unit plane oder auch chromaticity diagram genannt) wird durch die Punkte ![]() ,
, ![]() und
und ![]() aufgespannt, so daß sie sich auch als R + G + B = 1 definieren läßt. Der Farbvektor S einer beliebigen Farbe (bzw. dessen Verlängerung) schneidet die Einheitsebene in einem Punkt, der als Farbort oder chromaticity point der Farbe bezeichnet wird. Dieser Punkt besitzt die drei Farbwerte R, G und B bzw. die Farbwertanteile r, g und b, die so normiert sind, daß ihre Summe Eins ergibt.
aufgespannt, so daß sie sich auch als R + G + B = 1 definieren läßt. Der Farbvektor S einer beliebigen Farbe (bzw. dessen Verlängerung) schneidet die Einheitsebene in einem Punkt, der als Farbort oder chromaticity point der Farbe bezeichnet wird. Dieser Punkt besitzt die drei Farbwerte R, G und B bzw. die Farbwertanteile r, g und b, die so normiert sind, daß ihre Summe Eins ergibt.
Auch für außerhalb des Farbkegels liegende Vektoren, die Richter (1981) virtuelle Farbvalenzen nennt, können Farbgleichungen konstruiert werden, die sich aber nicht in einem Farbmisch-Experiment realisieren lassen. Das 1931 von der CIE eingeführte Normfarbsystem basiert auf drei derartigen Primärvalenzen ![]() ,
, ![]() und
und ![]() , deren als Normfarbwerte bezeichnete Farbwerte X, Y, Z nur positive Werte annehmen können. Die Normfarbwertanteile x, y, z werden dabei wieder so normiert, daß x + y + z = 1 gilt.
, deren als Normfarbwerte bezeichnete Farbwerte X, Y, Z nur positive Werte annehmen können. Die Normfarbwertanteile x, y, z werden dabei wieder so normiert, daß x + y + z = 1 gilt.
Smith und Pokorny (1996) nehmen einen linearen Zusammenhang zwischen den Farbwerten im ![]() -System und den spektralen Empfindlichkeiten der drei Zapfentypen an. Die Zapfenerregungen der Zapfentypen L, M und S lassen sich aus den Normfarbwerten X, Y und Z nach der sogenannten Hunt-Pointer-Estevez-Transformation (siehe z.B. Hunt, 1995; Fairchild, 1996) für Y = 100 folgendermaßen berechnen:
-System und den spektralen Empfindlichkeiten der drei Zapfentypen an. Die Zapfenerregungen der Zapfentypen L, M und S lassen sich aus den Normfarbwerten X, Y und Z nach der sogenannten Hunt-Pointer-Estevez-Transformation (siehe z.B. Hunt, 1995; Fairchild, 1996) für Y = 100 folgendermaßen berechnen:
Diese Transformationsmatrix ist so normiert, daß für das energiegleiche Spektrum L = M = S = 100 gilt. Umgekehrt lassen sich auch die Normfarbwerte (X,Y,Z) durch Multiplikation der Rezeptorempfindlichkeiten (L,M,S) mit der Inversen der Transformationsmatrix aus Gleichung 1.1 bestimmen.
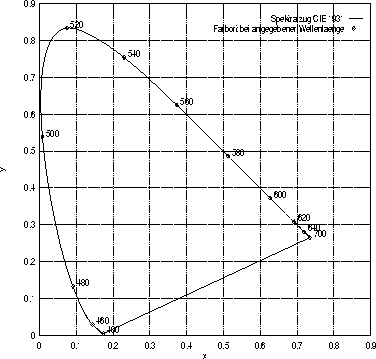
Die Normfarbtafel: In dieser Abbildung sind die Farborte der monochromatischen Reize der Wellenlängen zwischen 400 und 780 nm als hufeisenförmige Linie eingetragen.
Die als Normfarbtafel oder auch CIE-1931-Farbtafel bezeichnete Einheitsebene, die in Abbildung 1.2 zu sehen ist, wird so gewählt, daß bei ![]() ein rechter Winkel vorliegt. Trägt man in diese Normfarbtafel die Farborte
ein rechter Winkel vorliegt. Trägt man in diese Normfarbtafel die Farborte![]() der monochromatischen Reize von 400 bis 700 nm ein, erhält man den parabelförmigen Spektralzug (spectrum locus), der in Abbildung 1.2 ebenfalls eingezeichnet ist. Die gerade Verbindungslinie zwischen den beiden Endpunkten wird wegen des Farbtons der Farben auf dieser Linie auch als die Purpurlinie (purple line) bezeichnet. Außerhalb dieses parabelförmigen Gebildes liegen die virtuellen Farbvalenzen, denen kein physikalisch realisierbarer Reiz entspricht. Die Normfarbwertanteile
der monochromatischen Reize von 400 bis 700 nm ein, erhält man den parabelförmigen Spektralzug (spectrum locus), der in Abbildung 1.2 ebenfalls eingezeichnet ist. Die gerade Verbindungslinie zwischen den beiden Endpunkten wird wegen des Farbtons der Farben auf dieser Linie auch als die Purpurlinie (purple line) bezeichnet. Außerhalb dieses parabelförmigen Gebildes liegen die virtuellen Farbvalenzen, denen kein physikalisch realisierbarer Reiz entspricht. Die Normfarbwertanteile ![]() kennzeichnen den Punkt des energiegleichen Spektrums (equal-energy-stimulus), der einen normalerweise als farblos bzw. weiß wahrgenommenen Reiz repräsentiert. Die Darstellung von Farben in der Normfarbtafel wird sehr häufig zu deren Charakterisierung eingesetzt.
kennzeichnen den Punkt des energiegleichen Spektrums (equal-energy-stimulus), der einen normalerweise als farblos bzw. weiß wahrgenommenen Reiz repräsentiert. Die Darstellung von Farben in der Normfarbtafel wird sehr häufig zu deren Charakterisierung eingesetzt.
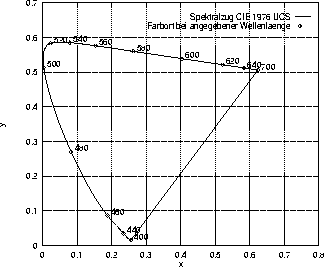
Allerdings entsprechen die euklidischen Abstände zwischen Farborten in der Normfarbtafel nicht den wahrnehmbaren Unterschieden zwischen den dazugehörigen Farben. Deshalb werden verschiedene Versuche unternommen, die Normfarbtafel so zu transformieren, daß die Abstände der transformierten Farbtafel eine Metrik![]() der Farbunterschiede bilden. Solche Farb-Systeme bezeichnet Richter (1981) als gleichabständige Farbart-Tafel oder Uniform-Chromaticity-Scale Diagram (kurz CIE-UCS-Farbtafel) und nennt als häufig in der Farbmetrik eingesetztes Beispiel die in Abbildung 1.3 dargestellte CIE-1976-Farbtafel. Sie basiert auf leicht modifizierten Daten von MacAdam (1937) und sieht folgende projektive Transformation der Normfarbtafel vor:
der Farbunterschiede bilden. Solche Farb-Systeme bezeichnet Richter (1981) als gleichabständige Farbart-Tafel oder Uniform-Chromaticity-Scale Diagram (kurz CIE-UCS-Farbtafel) und nennt als häufig in der Farbmetrik eingesetztes Beispiel die in Abbildung 1.3 dargestellte CIE-1976-Farbtafel. Sie basiert auf leicht modifizierten Daten von MacAdam (1937) und sieht folgende projektive Transformation der Normfarbtafel vor:
Die CIE-1976-UCS-Farbtafel: In dieser Abbildung sind die Farborte der monochromatischen Reize der Wellenlängen zwischen 400 und 780 nm als hufeisenförmige Linie eingetragen. Im Gegensatz zur Normfarbtafel ist diese Farbtafel annähernd längentreu, wie im Text näher erläutert wird.
![]()
Bei der Konstruktion solcher Farbtafeln werden die für die Transformation notwendigen Koeffizienten nicht aufgrund theoretischer Überlegungen festgesetzt, sondern müssen durch Ausprobieren gefunden werden. Auf diese Weise lassen sich nur approximativ gleichabständige Farb-Systeme finden und MacAdam (1942a) weist zudem darauf hin, daß sich prinzipiell keine projektive Transformation finden läßt, die zu einer UCS-Farbtafel führt. Die Verwendung einer anderen nicht-linearen Transformation könnte zwar zum gewünschten Ergebnis führen, würde aber auch bewirken, daß beispielsweise die geometrische Deutung der Graßmannschen Gesetze ihre Gültigkeit verlieren würde.
Ein anderer Ansatz zur Konstruktion eines Farbraums, in dem Entfernungen auch wahrnehmbaren Unterschieden entsprechen, besteht in der Festlegung des CIE-1976-![]() -Raumes (siehe Wyszecki & Stiles, 1982, S. 166 ff.), der oft zur Beschreibung von Farbdifferenzen eingesetzt wird. Dieser Farbraum besitzt die drei Dimensionen
-Raumes (siehe Wyszecki & Stiles, 1982, S. 166 ff.), der oft zur Beschreibung von Farbdifferenzen eingesetzt wird. Dieser Farbraum besitzt die drei Dimensionen ![]() und
und ![]() , die sich aus den Normfarbwerten X, Y, Z folgendermaßen berechnen lassen:
, die sich aus den Normfarbwerten X, Y, Z folgendermaßen berechnen lassen:
wobei gelten muß ![]() . Die Werte
. Die Werte ![]() bezeichnen die Farbwerte des Weißpunktes, der sich normalerweise aus dem Strahlungsspektrum der verwendeten Beleuchtung oder des Umfeldes ergibt. Üblicherweise wird
bezeichnen die Farbwerte des Weißpunktes, der sich normalerweise aus dem Strahlungsspektrum der verwendeten Beleuchtung oder des Umfeldes ergibt. Üblicherweise wird ![]() auf den Wert 100 gesetzt.
auf den Wert 100 gesetzt.
In diesem Zusammenhang repräsentiert dann ![]() die relative Helligkeit (lightness) des Reizes,
die relative Helligkeit (lightness) des Reizes, ![]() repräsentiert ungefähr den Wert bezüglich der Rot-Grünheit und
repräsentiert ungefähr den Wert bezüglich der Rot-Grünheit und ![]() den Wert bezüglich der Gelb-Blauheit (diese beiden Gegenfarben-Achsen sind in Abschnitt 1.2.2 eingehend dargestellt). Rechnet man diese Werte in zylindrische Koordinaten um, erhält man den Kennwerte
den Wert bezüglich der Gelb-Blauheit (diese beiden Gegenfarben-Achsen sind in Abschnitt 1.2.2 eingehend dargestellt). Rechnet man diese Werte in zylindrische Koordinaten um, erhält man den Kennwerte ![]() für die Buntheit (chroma) der Farbe und den in Grad angegebenen Farbwinkel
für die Buntheit (chroma) der Farbe und den in Grad angegebenen Farbwinkel ![]() zur numerischen Kennzeichnung des Farbtons (hue):
zur numerischen Kennzeichnung des Farbtons (hue):
Der CIE-1976-![]() -Raum hat nicht nur den Vorzug der Gleichabständigkeit, sondern er bietet auch die Möglichkeit, das Umfeld eines Reizes bzw. dessen Beleuchtung zu berücksichtigen.
-Raum hat nicht nur den Vorzug der Gleichabständigkeit, sondern er bietet auch die Möglichkeit, das Umfeld eines Reizes bzw. dessen Beleuchtung zu berücksichtigen.
Die in den letzten Abschnitten beschriebenen Farbräume können mehr oder weniger erfolgreich dazu eingesetzt werden, Unterschiede zwischen Farben durch deren euklidische Distanz innerhalb dieser Räume zu repräsentieren. Solche Maße für Farbdifferenzen sind auch zur Beurteilung von Kontext- und Adaptationseffekten wichtig.
Farbdifferenzen lassen sich auf verschiedene Arten erheben: Oft wird einfach die euklidische Distanz zwischen den zu vergleichenden Farben im Raum der xyL-Koordinaten, also der Normfarbwertanteile x und y sowie der Leuchtdichte L, berechnet (beispielsweise von Lucassen & Walraven, 1993). Unter der zuvor beschriebenen Annahme, daß der Farbwahrnehmung drei Systeme zugrunde liegen, deren Output linear mit den Normfarbwerten der entsprechenden Farbe zusammenhängt, erscheint es jedoch sinnvoller, als Abstandsmaß zweier Farben die euklidische Distanz aus deren Normfarbwerten zu errechnen:
wobei ![]() die Differenz der X-Koordinaten der beiden Farben bezeichnet,
die Differenz der X-Koordinaten der beiden Farben bezeichnet, ![]() die Differenz der Y-Koordinaten und
die Differenz der Y-Koordinaten und ![]() die Differenz der Z-Koordinaten. Dieses Abstandsmaß wird beispielsweise oft angewandt, um empirisch erhobene Einstellungen mit den von einem zu testenden Modell vorhergesagten zu vergleichen. Die Anwendung dieser Formel ist jedoch nicht unproblematisch, da der
die Differenz der Z-Koordinaten. Dieses Abstandsmaß wird beispielsweise oft angewandt, um empirisch erhobene Einstellungen mit den von einem zu testenden Modell vorhergesagten zu vergleichen. Die Anwendung dieser Formel ist jedoch nicht unproblematisch, da der ![]() -Raum nicht längentreu ist (siehe Abschnitt 1.1.3), die Gleichung 1.7 Abstände in diesem Raum aber immer gleich gewichtet.
-Raum nicht längentreu ist (siehe Abschnitt 1.1.3), die Gleichung 1.7 Abstände in diesem Raum aber immer gleich gewichtet.
Indow (1988) zeigt, daß für geringe Farbunterschiede, die nicht deutlich über einem ebenmerklichen Unterschied liegen, die Verwendung der folgenden Metrik unkritisch ist, die auf Nickerson (1936) zurückgeht:
![]()
Diesem Abstandsmaß liegt eine Repräsentation der zu vergleichenden Farben j und k durch ihre Munsell-Neuwerte Buntheit C (chroma), Buntton H (hue) und Helligkeitswert V (value) zugrunde, die aber nur bei Verwendung von Proben aus dem Munsell Book of Color problemlos gelingt.
Auf dem annähernd gleichabständigen CIE 1976 ![]() -Raum basiert das auch für größere Farbunterschiede geeignete Maß
-Raum basiert das auch für größere Farbunterschiede geeignete Maß ![]() (siehe z.B. Wyszecki & Stiles, 1982) für den Gesamtunterschied zweier Farben:
(siehe z.B. Wyszecki & Stiles, 1982) für den Gesamtunterschied zweier Farben:
![]()
wobei das Symbol ![]() den Unterschied der beiden zu vergleichenden Farben in der jeweiligen Dimension bezeichnet und
den Unterschied der beiden zu vergleichenden Farben in der jeweiligen Dimension bezeichnet und ![]() ,
, ![]() und
und ![]() wie in den Gleichungen 1.2 bis 1.4 beschrieben berechnet werden. Diese Formel, auch als CIE-1976-
wie in den Gleichungen 1.2 bis 1.4 beschrieben berechnet werden. Diese Formel, auch als CIE-1976-![]() -Farbunterschiedsformel (color-difference formula) bezeichnet, gilt allerdings nur unter bestimmten standardisierten Beobachtungsbedingungen.
-Farbunterschiedsformel (color-difference formula) bezeichnet, gilt allerdings nur unter bestimmten standardisierten Beobachtungsbedingungen.
Eine weitere Verbesserung dieser Formel (siehe Alman, 1993) stellen die daraus entwickelten sogenannten CIE ![]() -Gleichungen (CIE, 1995) dar, bei denen insbesondere die Gleichabständigkeit noch besser gewährleistet ist und die mit verschiedenen Parametern an die jeweilige Beobachtungssituation angepaßt werden können:
-Gleichungen (CIE, 1995) dar, bei denen insbesondere die Gleichabständigkeit noch besser gewährleistet ist und die mit verschiedenen Parametern an die jeweilige Beobachtungssituation angepaßt werden können:
Hierbei berechnet sich ![]() als Differenz der beiden nach Gleichung 1.6 berechneten Helligkeitswerte
als Differenz der beiden nach Gleichung 1.6 berechneten Helligkeitswerte ![]() der zu vergleichenden Farben,
der zu vergleichenden Farben, ![]() bezeichnet die Differenz der beiden Farbigkeitswerte
bezeichnet die Differenz der beiden Farbigkeitswerte ![]() , die nach Gleichung 1.5 berechnet wird, und
, die nach Gleichung 1.5 berechnet wird, und ![]() berechnet sich für kleine Farbdifferenzen nach
berechnet sich für kleine Farbdifferenzen nach
![]()
aus den beiden Farbtonwerte ![]() nach Gleichung 1.6. Für
nach Gleichung 1.6. Für ![]() kann man das geometrische Mittel der beiden
kann man das geometrische Mittel der beiden ![]() -Werte einsetzen. Die Parameter
-Werte einsetzen. Die Parameter ![]() und
und ![]() ermöglichen die Korrektur des Einflusses von solchen Störvariablen auf den Farbunterschied, die sich aufgrund des experimentellen Aufbaus ergeben können. Dabei handelt es sich um Variablen wie Reizgröße, Struktur der Reizoberfläche oder räumliche Anordnung der zu vergleichenden Reize. Unter den folgenden Referenz-Bedingungen der CIE (1995) werden diese drei Werte gleich Eins gesetzt: Beleuchtung durch die Lichtart D
ermöglichen die Korrektur des Einflusses von solchen Störvariablen auf den Farbunterschied, die sich aufgrund des experimentellen Aufbaus ergeben können. Dabei handelt es sich um Variablen wie Reizgröße, Struktur der Reizoberfläche oder räumliche Anordnung der zu vergleichenden Reize. Unter den folgenden Referenz-Bedingungen der CIE (1995) werden diese drei Werte gleich Eins gesetzt: Beleuchtung durch die Lichtart D![]() , Beleuchtungsstärke 1000 lux, Beobachter farbnormalsichtig, Beobachtungsart im Objektmodus, Reizgröße mehr als
, Beleuchtungsstärke 1000 lux, Beobachter farbnormalsichtig, Beobachtungsart im Objektmodus, Reizgröße mehr als ![]() Sehwinkel, Reizverteilung bei minimalem Abstand und direkt aneinandergrenzenden Begrenzungskanten, Größe der Farbunterschiede zwischen 0 und 5 CIE-
Sehwinkel, Reizverteilung bei minimalem Abstand und direkt aneinandergrenzenden Begrenzungskanten, Größe der Farbunterschiede zwischen 0 und 5 CIE-![]() -Einheiten und Reizstruktur homogen.
-Einheiten und Reizstruktur homogen.
Von den hier besprochenen Maßen erfaßt die ![]() -Differenz die wahrgenommenen Gesamt-Unterschiede zwischen zwei Farben am genauesten. Nun soll die Diskriminationsleistung für einzelne Farbattribute dargestellt werden.
-Differenz die wahrgenommenen Gesamt-Unterschiede zwischen zwei Farben am genauesten. Nun soll die Diskriminationsleistung für einzelne Farbattribute dargestellt werden.
Die Schwelle für Unterschiede im Farbton hängt von der jeweiligen Farbe ab, wie beispielsweise von Bedford und Wyszecki (1958) gezeigt wird: Sie liegt insbesondere in den Randbereichen des sichtbaren Spektrums wesentlich höher als im mittleren Bereich, außerdem nehmen die Unterschiedsschwellen mit der Reizintensität und der Reizgröße ab. Im Bereich der höchsten Empfindlichkeit bei ca. 590 nm lassen sich unter geeigneten Bedingungen monochromatische Reize unterscheiden, deren Wellenlängenmaximum nur 0.2 nm auseinanderliegt, wie LeGrand (1968) berichtet.
Auch für Unterschiede in der Sättigung zeigen sich Abhängigkeiten von der jeweiligen Farbe bzw. von deren bunttongleicher Wellenlänge.![]() Priest und Brickwedde (1938) untersuchen, wie viel monochromatisches Licht zu einem achromatischen Licht hinzugemischt werden muß, damit die Mischung nicht mehr als Weiß wahrgenommen wird. Sie kommen zu dem Ergebnis, daß die Größe eines ebenmerklichen Sättigungs-Unterschiedes systematisch von der Wellenlänge
Priest und Brickwedde (1938) untersuchen, wie viel monochromatisches Licht zu einem achromatischen Licht hinzugemischt werden muß, damit die Mischung nicht mehr als Weiß wahrgenommen wird. Sie kommen zu dem Ergebnis, daß die Größe eines ebenmerklichen Sättigungs-Unterschiedes systematisch von der Wellenlänge ![]() des präsentierten Spektralreizes abhängt: Für Reize, deren bunttongleiche Wellenlänge aus den Randbereichen des sichtbaren Spektrums stammt, ist die Unterschiedsschwelle relativ niedrig, während ihr Maximum in der Mitte des sichtbaren Bereichs bei etwa 570 nm liegt (siehe auch Wright & Pitt, 1937). Ähnliche Abhängigkeiten der Sättigung von der bunttongleichen Wellenlänge finden auch Jones und Lowry (1926) und Martin, Warburton und Morgan (1933), bei denen die minimale Anzahl an Unterscheidbarkeits-Schritten bei 570 nm liegt, während an den Enden des sichtbaren Spektrums drei mal so hohe Werte gefunden werden. Auch monochromatische Reize können sich also in ihrer Sättigung unterscheiden.
des präsentierten Spektralreizes abhängt: Für Reize, deren bunttongleiche Wellenlänge aus den Randbereichen des sichtbaren Spektrums stammt, ist die Unterschiedsschwelle relativ niedrig, während ihr Maximum in der Mitte des sichtbaren Bereichs bei etwa 570 nm liegt (siehe auch Wright & Pitt, 1937). Ähnliche Abhängigkeiten der Sättigung von der bunttongleichen Wellenlänge finden auch Jones und Lowry (1926) und Martin, Warburton und Morgan (1933), bei denen die minimale Anzahl an Unterscheidbarkeits-Schritten bei 570 nm liegt, während an den Enden des sichtbaren Spektrums drei mal so hohe Werte gefunden werden. Auch monochromatische Reize können sich also in ihrer Sättigung unterscheiden.
Betrachtet man umgekehrt, wieviel achromatisches Licht zu einem monochromatischem Reiz bestimmter Wellenlänge hinzugemischt werden muß, damit ein sichtbarer Unterschied zu einem rein monochromatischem Reiz gleicher Wellenlänge und Leuchtdichte auftritt, so zeigt sich, daß die Unterschiedsschwelle in dieser Richtung für alle Wellenlängen relativ konstant ist (siehe Wright & Pitt, 1935).
Besonders empfindlich ist das visuelle System für Unterschiede in der Helligkeit eines Reizes, wobei der gerade eben wahrnehmbare Leuchtdichte-Unterschied vom Ausgangsniveau der Leuchtdichte abhängt. Nach Lowry (1931) liegt im Bereich zwischen 80 und 300 ![]() die maximale Empfindlichkeit für Leuchtdichteunterschiede vor, bei Leuchtdichten über 100
die maximale Empfindlichkeit für Leuchtdichteunterschiede vor, bei Leuchtdichten über 100 ![]() trifft hierbei das Webersche Gesetz annähernd zu: Es besteht ein konstanter Zusammenhang zwischen dem Zuwachs an Leuchtdichte
trifft hierbei das Webersche Gesetz annähernd zu: Es besteht ein konstanter Zusammenhang zwischen dem Zuwachs an Leuchtdichte ![]() , der für einen wahrnehmbaren Unterschied erforderlich ist, und dem Ausgangsniveau der Leuchtdichte L. Im Bereich dieser Leuchtdichten liegt der Weber-Bruch
, der für einen wahrnehmbaren Unterschied erforderlich ist, und dem Ausgangsniveau der Leuchtdichte L. Im Bereich dieser Leuchtdichten liegt der Weber-Bruch ![]() bei etwa 0.01, es sind also Veränderungen der Leuchtdichte um etwa 1% wahrnehmbar.
bei etwa 0.01, es sind also Veränderungen der Leuchtdichte um etwa 1% wahrnehmbar.
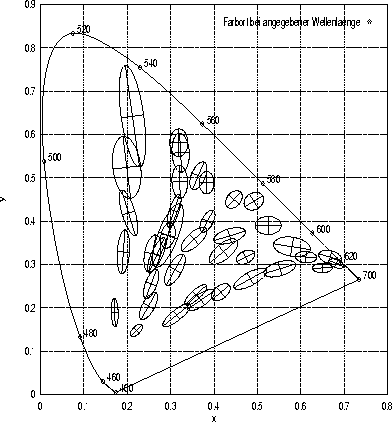
Nach der Darstellung der Unterschiedsschwellen des visuellen Systems für einzelne Farbattribute im letzten Abschnitt soll nun die Unterscheidbarkeit von Farben im dreidimensionalen Farbraum besprochen werden. Sie kann durch Analyse der Streuungen bestimmt werden, die bei wiederholten Abgleichen zu einem vorgegebenen Standardreiz auftreten: Diese Streuungen lassen sich durch Ellipsoide im dreidimensionalen Farbraum beschreiben, deren Zentroid beim Standardreiz liegt und deren Radien durch die Standardabweichung der Daten in die jeweilige Richtung gegeben sind. Solche Ellipsoide können als Kontur eines ebenmerklichen Unterschieds interpretiert werden und werden deshalb als Diskriminationsellipsoide bezeichnet. Eine Projektion solcher Ellipsoide auf die Normfarbtafel ist in Abbildung 1.4 zu sehen.
Voraussetzung für die Bestimmung der Diskriminationsellipsoide ist multivariate Normalverteilung der betreffenden Variable, von der nach Brown (1952a) bei den durch Farbabgleiche erhobenen Farborten auch ausgegangen werden kann (siehe auch Seite ![]() und Silberstein & MacAdam, 1945). Tatsuoka (1971) stellt die multivariate Normalverteilung und die Bestimmung der Diskriminationsellipsoide zu einer gegebenen oder zu schätzenden Verteilung dar. Die allgemeine Form der multivariaten Normalverteilung lautet im p-dimensionalen Fall für die Dichtefunktion
und Silberstein & MacAdam, 1945). Tatsuoka (1971) stellt die multivariate Normalverteilung und die Bestimmung der Diskriminationsellipsoide zu einer gegebenen oder zu schätzenden Verteilung dar. Die allgemeine Form der multivariaten Normalverteilung lautet im p-dimensionalen Fall für die Dichtefunktion ![]() eines Zufallsvektors
eines Zufallsvektors ![]()
![]()
wobei ![]() den Erwartungswertsvektor - auch Zentroid genannt - und
den Erwartungswertsvektor - auch Zentroid genannt - und ![]() die Varianz-Kovarianz-Matrix bezeichnen. Bei Betrachtung der Dichtefunktion der mehrdimensionalen Normalverteilung in Gleichung 1.12 erkennt man, daß die Daten
die Varianz-Kovarianz-Matrix bezeichnen. Bei Betrachtung der Dichtefunktion der mehrdimensionalen Normalverteilung in Gleichung 1.12 erkennt man, daß die Daten ![]() nur in der quadratischen Form im Exponenten der Dichtefunktion vorkommen. Daraus ergibt sich, daß für jede Konstante
nur in der quadratischen Form im Exponenten der Dichtefunktion vorkommen. Daraus ergibt sich, daß für jede Konstante ![]() aus
aus ![]() die Menge aller
die Menge aller ![]() , für die gilt
, für die gilt
![]()
auch denselben Wert von ![]() und somit auch dieselbe Wahrscheinlichkeitsdichte besitzt. Solche als Isodensiten bezeichneten Konturen gleicher Wahrscheinlichkeitsdichte umschließen eine Fläche bzw. einen Raum, der als Konfidenzintervall um den Mittelwertsvektor aufgefaßt werden kann. Der zu diesem Konfidenzintervall gehörende Wahrscheinlichkeitswert hängt dabei monoton von C ab. Oft ist die Dichtefunktion der Daten aber nicht a priori gegeben, so daß die Varianz-Kovarianz-Matrix aus den Daten folgendermaßen geschätzt werden muß:
und somit auch dieselbe Wahrscheinlichkeitsdichte besitzt. Solche als Isodensiten bezeichneten Konturen gleicher Wahrscheinlichkeitsdichte umschließen eine Fläche bzw. einen Raum, der als Konfidenzintervall um den Mittelwertsvektor aufgefaßt werden kann. Der zu diesem Konfidenzintervall gehörende Wahrscheinlichkeitswert hängt dabei monoton von C ab. Oft ist die Dichtefunktion der Daten aber nicht a priori gegeben, so daß die Varianz-Kovarianz-Matrix aus den Daten folgendermaßen geschätzt werden muß:
![]()
Man erhält im Fall von aus den Daten zu schätzender Varianz-Kovarianz-Matrix nach Fahrmeir und Hamerle (1996) die sogenannte Mahalanobis-Distanz
![]()
mittels derer man den ![]() -Vertrauensbereich um eine p-variate normalverteilt Zufallsvariable durch
-Vertrauensbereich um eine p-variate normalverteilt Zufallsvariable durch
![]()
bestimmen kann. Auch durch diese Mahalanobis-Distanz wird wieder ein Ellipsoid mit Mittelpunkt ![]() definiert.
definiert.
Die bekannten von MacAdam (1942b) bestimmten Diskriminationsellipsen kommen auf ganz andere Art als gerade beschrieben aus den Einstellungen von zwei Versuchspersonen zustande, die bei konstanter Leuchtdichte das Mischungsverhältnis zweier annähernd monochromatischer Lichtstrahlen so verändern, daß eine Farbgleichheit zwischen dieser Mischung und einem vorgegebenen Standard hergestellt wird. Die beiden zu mischenden Reize sind so ausgewählt, daß bei einem bestimmten Mischungsverhältnis Farbgleichheit zum vorgegebenen Standard möglich ist. Aus jeweils 50 solchen Einstellungen wird die Standardabweichung der Normfarbwertanteile bestimmt und auf die Verbindungsgerade zwischen den Farborten der beiden zu mischenden Reize in der Normfarbtafel eingetragen. Die bei verschiedenen Reizpaaren bestimmten Standardabweichungen um denselben Standardreiz werden von MacAdam (1942b) durch eine Ellipse beschrieben, die angibt, ab welchen Differenzen Farben unterschieden werden können. Man erhält auf diese Weise einen Überblick darüber, wie sich die Unterscheidbarkeit von Farben in verschiedenen Regionen der Normfarbtafel verändert.
Die gerade beschriebenen Diskriminationsellipsen von MacAdam (1942b) kommen unter der Einschränkung zustande, daß die Leuchtdichte konstant gehalten wird. Brown und MacAdam (1949) bestimmen die in Abbildung 1.4 gezeigten Diskriminationsellipsen durch Mischung von drei Primärreizen, wobei auch die Leuchtdichte der Mischung variieren kann. Aufgrund der hier angenommenen trivariaten Normalverteilung beträgt die Wahrscheinlichkeit dafür, daß ein Datenpunkt in das entsprechende Ellipsoid fällt, allerdings nur etwa 20%, wie Brown, Howe, Jackson und Morris (1956) nachweisen (und nicht 68%, wie Brown und MacAdam (1949) fälschlicherweise behaupten).
Diskriminationsellipsoide nach Brown und MacAdam (1949):
In dieser Abbildung sind 38 der von Brown und MacAdam (1949) bei der Versuchsperson WRJB erhobenen Diskriminationsellipsen (in zehnfacher Vergrößerung) sowie deren Hauptachsen zu sehen. Grundlage der Darstellung sind die von Brown und MacAdam (1949, S.824 f.) in ihrer Tabelle V veröffentlichten Schätzungen der Ellipsenparameter. Die monokular präsentierten, 2![]() großen Reize können von den Versuchspersonen auf allen drei Dimensionen verändert werden.
großen Reize können von den Versuchspersonen auf allen drei Dimensionen verändert werden.
Brown (1957) versucht schließlich, die genannten Ergebnisse noch zu weiter generalisieren, indem er ähnliche Untersuchungen an zwölf Versuchspersonen durchführt, von denen nur der Autor selbst mit der Fragestellung vertraut ist. Bei dieser Untersuchung kann der 10![]() große Zielreiz beidäugig betrachtet werden, wobei von jedem Beobachter für jeden der 22 Zielreize 60 Farbabgleiche durchgeführt werden. Auch die Ergebnisse auf dieser breiten empirischen Basis stimmen mit den beiden zuvor genannten in bezug auf die Größe und Lage der Diskriminationsellipsen überein.
große Zielreiz beidäugig betrachtet werden, wobei von jedem Beobachter für jeden der 22 Zielreize 60 Farbabgleiche durchgeführt werden. Auch die Ergebnisse auf dieser breiten empirischen Basis stimmen mit den beiden zuvor genannten in bezug auf die Größe und Lage der Diskriminationsellipsen überein.
Brown (1957) untersucht, wovon die Größe der Diskriminationsellipsoide neben dem Farbort noch abhängt. Er kann dabei Trainingseffekte bei den anfangs ungeübten Beobachtern nachweisen: In der ersten Sitzung unterscheiden sich die aus ihren Einstellungen bestimmten Ellipsen in der Normfarbtafel zwischen den Versuchspersonen deutlich in ihrer Größe und Orientierung, in der siebzehnten Sitzung ist dies nicht mehr der Fall. Brown (1957) interpretiert dieses Ergebnis so, ``that all of the observers by the time they had matched the seventeenth color center could be considered skilled observers'' (Brown, 1957, S.140). Neben der größeren Übereinstimmung zwischen den Ellipsen fällt außerdem auf, daß die durchschnittliche Größe der Ellipsen deutlich abnimmt.
Nach Brown (1952b) hängt die Größe der Diskriminationsellipsen auch von der Farbe eines gleichzeitig dargebotenen Umfeldes ab: Die Diskriminationsleistung der Versuchsperson ist am höchsten, wenn der Reiz vor einem Hintergrund dargeboten wird, dessen Farbe dem Reiz selbst ähnlich ist. Eine Vergrößerung des Reizes von 2![]() auf 12
auf 12![]() bringt außerdem für alle Hintergrundvarianten eine deutliche Verbesserung der Diskriminiationsfähigkeit, da sich in diesem Fall auch andersfarbige Hintergrundbeleuchtungen nicht so stark auswirken wie bei kleineren Zielreizen.
bringt außerdem für alle Hintergrundvarianten eine deutliche Verbesserung der Diskriminiationsfähigkeit, da sich in diesem Fall auch andersfarbige Hintergrundbeleuchtungen nicht so stark auswirken wie bei kleineren Zielreizen.